Исследователи из Университета Сайтама и Национального института фундаментальной биологии в Японии идентифицировали ключевой компонент, отвечающий за реакцию венериной мухоловки на прикосновение. Им оказался специфический ионный канал, названный DmMSL10. Этот канал функционирует как основной механосенсор, преобразуя слабое физическое воздействие в мощный электрический сигнал, который заставляет ловушку захлопываться. Результаты работы опубликованы в журнале Nature Communications.
Ловушка хищного растения Dionaea muscipula приводится в действие высокочувствительными сенсорными волосками. Для срабатывания механизма требуется два последовательных прикосновения к этим волоскам. Такая система позволяет растению отличать потенциальную добычу от ложных раздражителей, например, капель дождя, и избегать бесполезного расхода энергии.
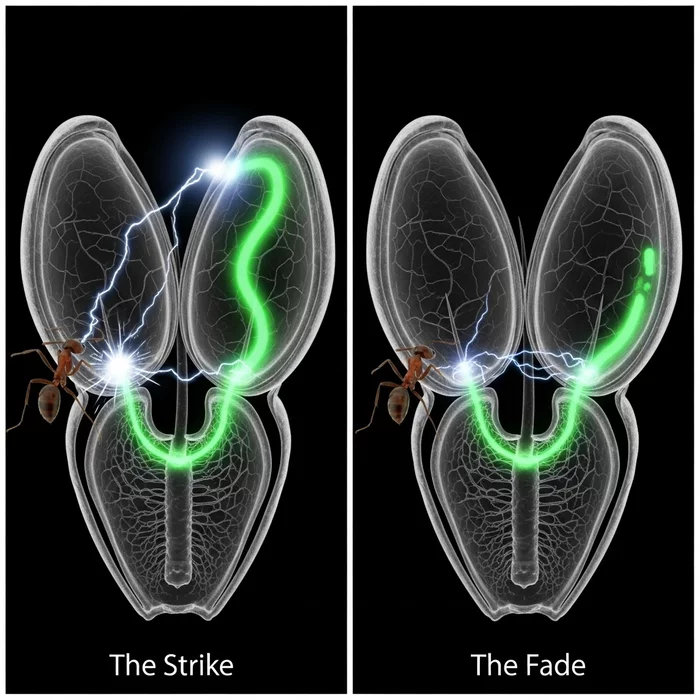
На молекулярном уровне процесс начинается с небольшого локального изменения электрического заряда у основания сенсорного волоска при его легком изгибе. Более сильный изгиб вызывает более значительный первоначальный электрический отклик. Когда этот сигнал достигает определенного порогового значения, он запускает мощный электрический всплеск, действующий по принципу «все или ничего», подобно щелчку выключателя.
Ключевая роль в этом процессе принадлежит ионному каналу DmMSL10. Он действует как усилитель. Канал многократно увеличивает слабый электрический сигнал от первоначального прикосновения до тех пор, пока его сила не превысит порог, необходимый для активации ловушки. Этот механизм напоминает работу нервной системы животных, хотя у растений нервы отсутствуют.
После преодоления порога генерируется не только основной электрический всплеск, но и химический сигнал в виде волны ионов кальция (Ca2+). Этот комбинированный электрохимический сигнал распространяется от основания сенсорных волосков к створкам листа, передавая команду на закрытие.
Для подтверждения своих выводов команда под руководством Масацугу Тойоты провела серию экспериментов. Сначала они спроектировали венерины мухоловки, экспрессирующие белок GCaMP6f. Это позволило визуально наблюдать за химическими и электрическими изменениями в живом растении в момент прикосновения к сенсорному волоску. Исследователи смогли увидеть как локальный заряд от слабого касания, так и мощный всплеск и волну Ca2+ от сильного.
Решающим доказательством стала генная инженерия растения, у которого отсутствовал ионный канал DmMSL10. Такие модифицированные экземпляры демонстрировали значительно более слабую реакцию на стимулы, что подтвердило критическую роль DmMSL10 в усилении сигнала.
Чтобы проверить выводы в условиях, имитирующих естественную среду, ученые создали в лаборатории небольшую экосистему. В ней муравьи могли свободно перемещаться как по обычным венериным мухоловкам (с DmMSL10), так и по генетически измененным (без DmMSL10). Растения с каналом DmMSL10 закрывали свои ловушки на муравьях значительно чаще, а их сенсорные волоски изгибались активнее. Модифицированные растения практически не реагировали на насекомых.
По словам соавтора исследования, биолога растений Хираку Суды, эти открытия имеют более широкое значение. Они предполагают, что молекулярные механизмы механосенсорики — чувства осязания у растений — могут быть общими для многих видов, а не только для венериной мухоловки, эндемика Северной и Южной Каролины в США.
Изображение носит иллюстративный характер
Ловушка хищного растения Dionaea muscipula приводится в действие высокочувствительными сенсорными волосками. Для срабатывания механизма требуется два последовательных прикосновения к этим волоскам. Такая система позволяет растению отличать потенциальную добычу от ложных раздражителей, например, капель дождя, и избегать бесполезного расхода энергии.
На молекулярном уровне процесс начинается с небольшого локального изменения электрического заряда у основания сенсорного волоска при его легком изгибе. Более сильный изгиб вызывает более значительный первоначальный электрический отклик. Когда этот сигнал достигает определенного порогового значения, он запускает мощный электрический всплеск, действующий по принципу «все или ничего», подобно щелчку выключателя.
Ключевая роль в этом процессе принадлежит ионному каналу DmMSL10. Он действует как усилитель. Канал многократно увеличивает слабый электрический сигнал от первоначального прикосновения до тех пор, пока его сила не превысит порог, необходимый для активации ловушки. Этот механизм напоминает работу нервной системы животных, хотя у растений нервы отсутствуют.
После преодоления порога генерируется не только основной электрический всплеск, но и химический сигнал в виде волны ионов кальция (Ca2+). Этот комбинированный электрохимический сигнал распространяется от основания сенсорных волосков к створкам листа, передавая команду на закрытие.
Для подтверждения своих выводов команда под руководством Масацугу Тойоты провела серию экспериментов. Сначала они спроектировали венерины мухоловки, экспрессирующие белок GCaMP6f. Это позволило визуально наблюдать за химическими и электрическими изменениями в живом растении в момент прикосновения к сенсорному волоску. Исследователи смогли увидеть как локальный заряд от слабого касания, так и мощный всплеск и волну Ca2+ от сильного.
Решающим доказательством стала генная инженерия растения, у которого отсутствовал ионный канал DmMSL10. Такие модифицированные экземпляры демонстрировали значительно более слабую реакцию на стимулы, что подтвердило критическую роль DmMSL10 в усилении сигнала.
Чтобы проверить выводы в условиях, имитирующих естественную среду, ученые создали в лаборатории небольшую экосистему. В ней муравьи могли свободно перемещаться как по обычным венериным мухоловкам (с DmMSL10), так и по генетически измененным (без DmMSL10). Растения с каналом DmMSL10 закрывали свои ловушки на муравьях значительно чаще, а их сенсорные волоски изгибались активнее. Модифицированные растения практически не реагировали на насекомых.
По словам соавтора исследования, биолога растений Хираку Суды, эти открытия имеют более широкое значение. Они предполагают, что молекулярные механизмы механосенсорики — чувства осязания у растений — могут быть общими для многих видов, а не только для венериной мухоловки, эндемика Северной и Южной Каролины в США.











